Что такое интуиция на самом деле? Первая карта мозга показывает, как работает чутьё
Знакомо чувство, когда вы, не успев подумать, делаете выбор, который кажется единственно верным? Или когда внутренний голос настойчиво шепчет: «Сюжет повернётся именно так». Мы называем это интуицией, шестым чувством, чутьём. Долгое время это явление обитало на границе психологии и мистики. Но что, если интуиция — это не загадочный дар, а отточенный до совершенства механизм работы нашего мозга?
Недавнее исследование, ставшее плодом беспрецедентного сотрудничества двенадцати лабораторий, впервые позволило создать полную карту мозговой активности млекопитающего в момент принятия решения. И результаты оказались куда более ошеломляющими, чем просто красивая картинка работающих нейронов. Они заставляют нас по-новому взглянуть на то, как мы думаем, как воспринимаем реальность и что на самом деле скрывается за нашими «спонтанными» озарениями.
«ЦЕРН для мозга»: как заглянуть в мысли мыши
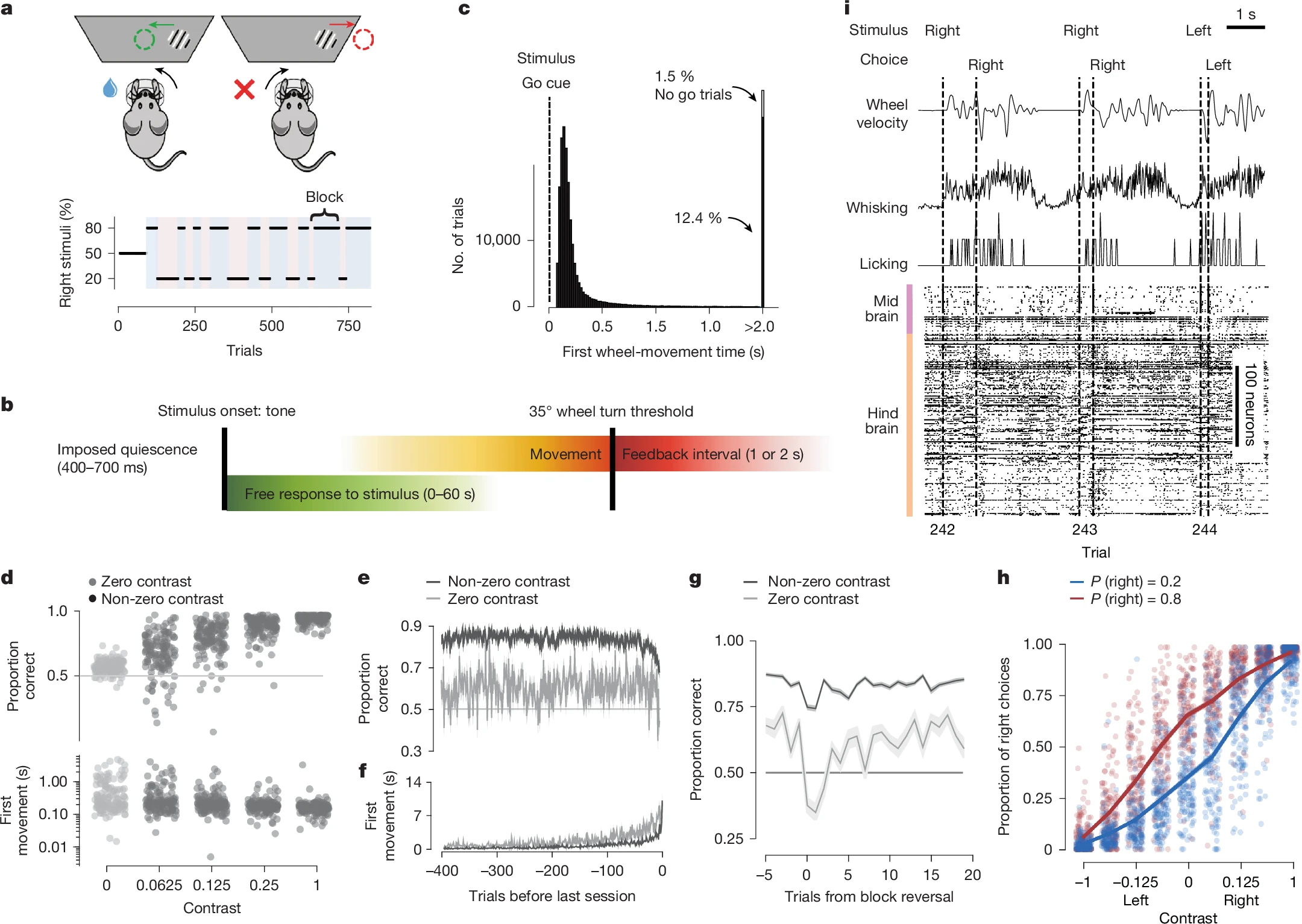
Прежде всего, стоит осознать масштаб проделанной работы. Десятилетиями нейробиология сталкивалась с фундаментальным ограничением: заглянуть в мозг можно было лишь «через замочную скважину». Один электрод регистрирует активность лишь небольшой группы нейронов, а превратить живое существо в «подушечку для иголок» невозможно. Чтобы обойти эту проблему, учёные пошли по пути физиков-ядерщиков, создавших ЦЕРН. Они объединили усилия, создав своего рода «ЦЕРН для нейробиологии».
В каждой из двенадцати лабораторий мыши выполняли одну и ту же задачу: с помощью крошечного руля из Lego они должны были переместить полосатую мишень на экране. Сложность заключалась в том, что контрастность мишени менялась. Иногда она была видна чётко, а иногда практически сливалась с фоном, заставляя животное полагаться не на зрение, а на свой предыдущий опыт. Экспериментаторы намеренно создавали «предвзятость», заставляя мишень чаще появляться с одной стороны экрана.
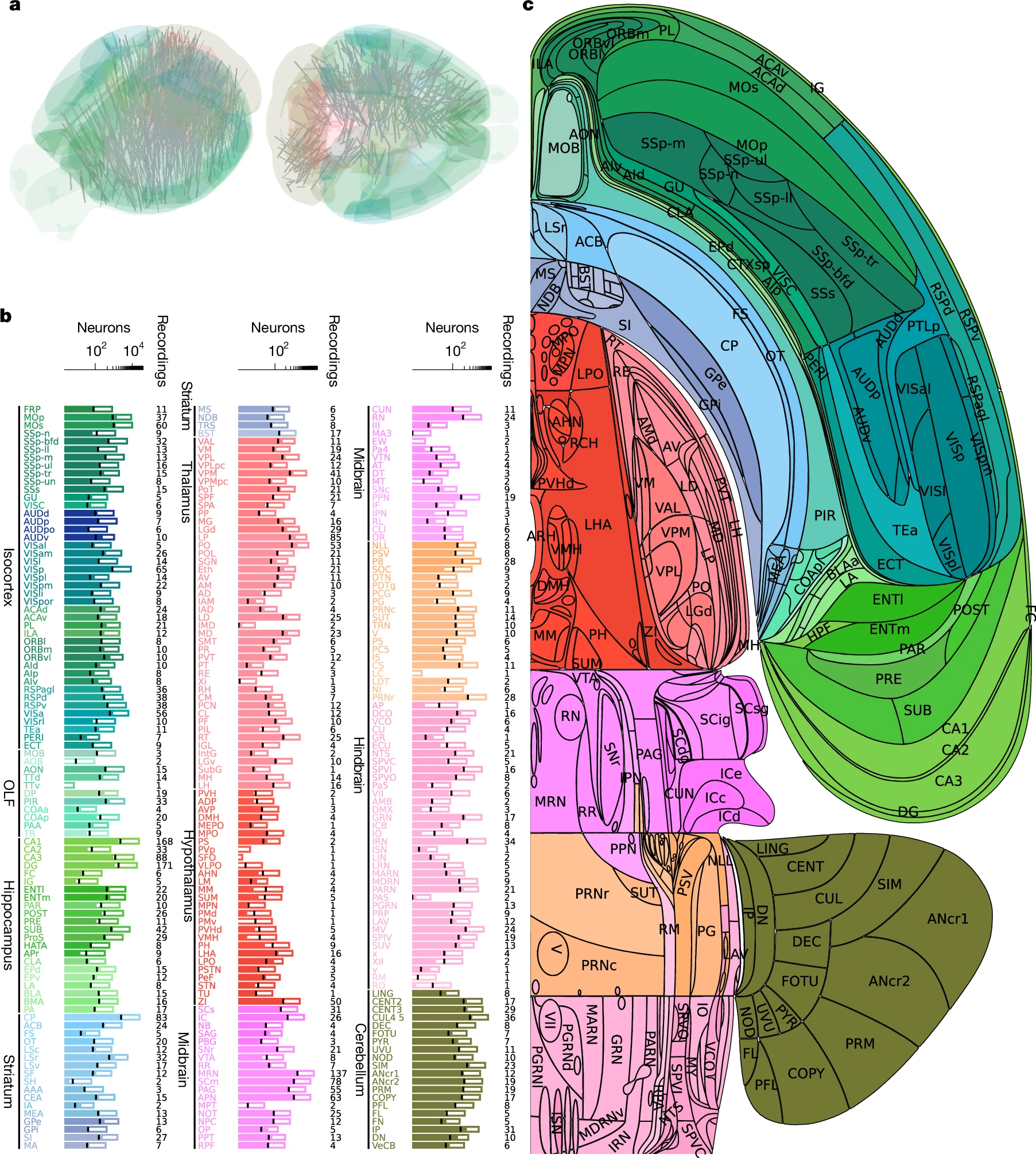
Каждая лаборатория регистрировала активность в своей зоне мозга, но с перекрытиями для калибровки данных. В итоге, собрав воедино мозаику из более чем 650 000 нейронов, учёные получили то, чего не видел ещё никто — целостную динамическую картину сложнейшего когнитивного процесса.
Решение — это не точка, а процесс
Первый миф, который развеяло исследование, — это идея о существовании некоего «центра принятия решений». Мы привыкли думать, что где-то в префронтальной коре есть условная «красная кнопка», на которую нажимает наш внутренний генеральный директор. Реальность оказалась сложнее и элегантнее.
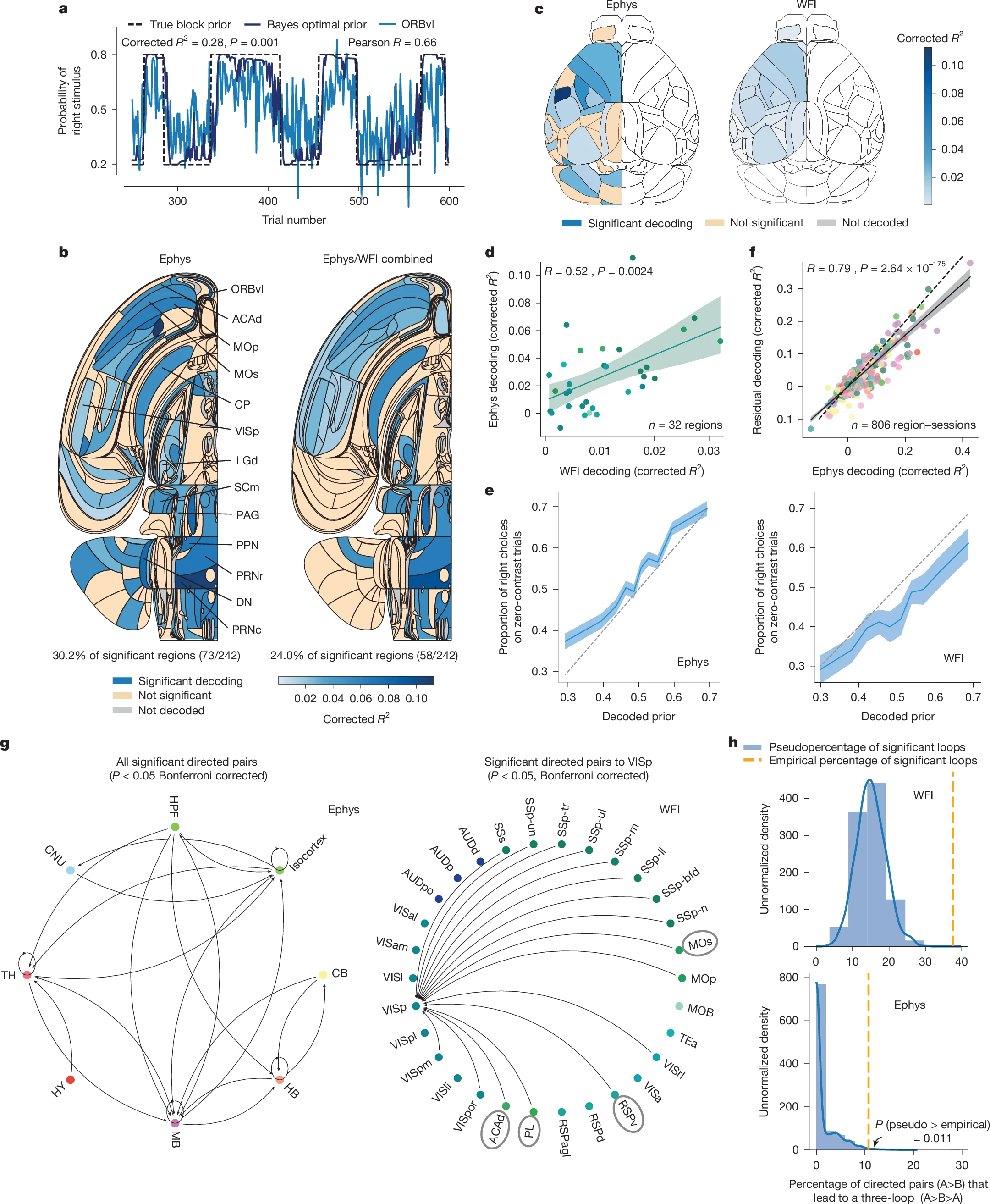
Карта активности показала: в принятии решения участвуют десятки областей мозга. Это не команда одного начальника, а скорее гул целого парламента, где разные фракции (зоны мозга) взвешивают аргументы и постепенно приходят к консенсусу. Процесс не локализован, он распределён по всей нейронной сети.
Более того, «решение» начинает формироваться задолго до того, как мышь повернёт руль. Сигналы, соответствующие будущему выбору, нарастали в мозге ещё в состоянии покоя, до появления стимула. Они постепенно накапливались, словно вода в плотине, и лишь по достижении определённого порога приводили к физическому действию. Это фундаментально меняет наше представление о выборе: он не происходит в один момент, а вызревает в глубинах нейронных сетей.
Предвзятый мозг: как опыт формирует реальность
Но самое поразительное открытие ждало исследователей на самом «входе» информации. Оказалось, что наши ожидания и прошлый опыт влияют на восприятие ещё до того, как мы успеваем что-либо осознать.
Сигналы от глаз мыши, едва начав свой путь в таламус — своего рода «диспетчерскую» мозга, — уже были изменены! Мозг, основываясь на опыте (например, «в последнее время мишень чаще появляется слева»), заранее «подкрашивал» входящую зрительную информацию. Он не просто пассивно получал картинку мира, а активно интерпретировал её, исходя из своих убеждений.
Именно здесь, по мнению учёных, и может скрываться нейробиологическая основа интуиции. То, что мы ощущаем как «внутреннее знание» или «предчувствие», на самом деле — результат молниеносной и неосознанной обработки данных, уже искажённых нашим прошлым опытом. Наш мозг — это машина для предсказаний. Он постоянно строит модель мира и, сталкиваясь с неполной информацией (как в случае с низкоконтрастной мишенью), заполняет пробелы наиболее вероятным, с его точки зрения, сценарием. Этот процесс настолько быстр и глубок, что до нашего сознания доходит уже готовый результат — то самое «чутьё».
Как отмечает Лоуренс Хант из Оксфорда, особенно интригует то, что мозг формирует ожидания не только на основе объективных событий, но и на основе наших собственных прошлых выборов. Иными словами, мы видим то, что ожидаем увидеть, потому что мы привыкли так действовать. Наш субъективный опыт становится фильтром, через который мы смотрим на мир.
Свобода воли под вопросом?
Естественно, такие выводы не могли не затронуть вечный философский вопрос о свободе воли. Если мозг предопределяет реакцию ещё до осознанного выбора, и если наше восприятие заранее искажено опытом, свободны ли мы в своих решениях?
Александр Пуже, один из авторов работы, даёт довольно прямой ответ: мозг и окружающий мир — это детерминированная система. Физические законы, управляющие нейронами, неумолимы. Зная все начальные условия, можно с определённой долей вероятности предсказать будущее решение.
Однако это не повод для фатализма. Ключевое слово здесь — «окружающий мир». Он постоянно меняется, поставляя новую информацию. Эта непредсказуемость внешних событий не позволяет системе быть полностью замкнутой. Мы постоянно обновляем наши «внутренние карты» и корректируем ожидания. Таким образом, хотя работа мозга подчиняется строгим законам, его взаимодействие с хаотичной реальностью создаёт пространство для вариативности и того, что мы ощущаем как свободу выбора.
Это исследование — не просто шаг вперёд для нейробиологии. Это окно в самые основы нашего познания. Оно показывает, что наша реальность — это не пассивное отражение мира, а активный конструкт, создаваемый нашим мозгом на основе прошлого. А интуиция — не магия, а высшее проявление эффективности этого удивительного механизма. И понимание этого механизма в будущем может помочь в лечении таких состояний, как аутизм, при котором, как предполагается, нарушена именно способность гибко обновлять свои внутренние модели мира.











0 комментариев
Добавить комментарий
Добавить комментарий